

Русские из работы Густава-Теодора Паули «Народы России»
Русское население является довольно однородным в антропологическом отношени (1).
По трём антропологическим признакам (ширина головы, ширина носа, толщина губ) русские популяции достоверно не отличаются от западноевропейских. По остальным размерам головы и лица они близки к центральному европейскому варианту, характеризующемуся средними размерами. Средние величины группы или совпадают с центральными западноевропейскими величинами, или отклоняются от них, оставаясь, однако, в пределах колебаний западных групп (1).
Для русского населения характерно отсутствие эпикантуса. Из более чем 8,5 тысяч обследованных русских мужского пола эпикантус выявлен у 12 человек, к тому же только в зачаточном состоянии. Так же редок эпикантус у населения Центральной Европы (2).
Встречаемость прямого профиля носа у русских составляет 75 %, что выше среднего значения по Европе 70 % (3). Встречаемость вогнутого профиля носа у русских составляет 9 %, что приближается к средним значениям по Западной и Центральной Европе 10 % (3).
Сочетание признаков, характерных для русских популяций в сравнении с западноевропейским центральным вариантом (1):
- Сравнительно светлая пигментация. Светлых волос 33 %, волос среднего оттенка 53,2 %, тёмных 13,8 %. Светлых глаз 45 %, светло-смешанных 49,9 %, тёмных 5,1 % (1).
- Средний рост бровей и бороды;
- Умеренная ширина лица;
- Преобладание среднего горизонтального профиля и средневысокого переносья;
- Меньший наклон лба и более слабое развитие надбровья.
Результаты краниологических исследований также демонстрируют исключительно высокое морфологическое сходство. Все локальные варианты проявляются в пределах основного единого и гомогенного краниологического типа русских (4, 5).
Антропологическое зонирование
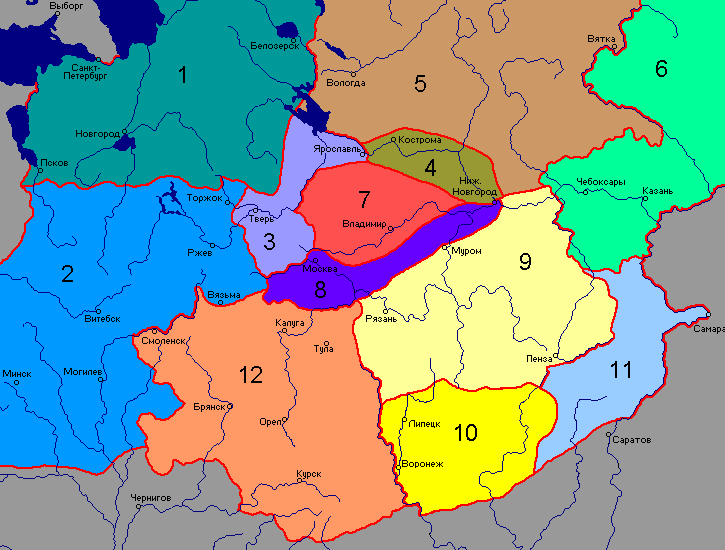
Географические зоны антропологических типов русских: 1 — ильменско-белозёрская; 2 — валдайская; 3 — западная верхневолжская; 4 — восточная верхневолжская; 5 — вологдо-вятская; 6 — вятско-камская; 7 — клязьминская; 8 — центральная; 9 — дон-сурская; 10 — степная; 11 — средневолжская; 12 — верхнеокская
Характер географической изменчивости антропологических признаков и их сочетания позволил выделить среди русского населения ряд локальных антропологических типов, получивших названия в соответствии с географической привязкой.
 Русская антропологическая экспедиция (1955—1959) — антропологическая экспедиция под руководством Бунака В. В. Целью экспедиции являлось изучение антропологических характеристик русского народа. Проводилась в период с 1955 по 1959 годы.
Русская антропологическая экспедиция (1955—1959) — антропологическая экспедиция под руководством Бунака В. В. Целью экспедиции являлось изучение антропологических характеристик русского народа. Проводилась в период с 1955 по 1959 годы.
Русская антропологическая экспедиция была организована Отделом антропологии Института этнографии АН СССР при участии НИИ и Музея антропологии МГУ. Научным руководителем экспедиции являлся антрополог Бунак В. В.
Исследования проводились в зоне расселения русского народа в XI—XIV веках, центральной части Русской равнины между Верхней Волгой и Окой — Ростово-Суздальское княжество, областях Московского, Рязанского, Смоленского, Тверского княжеств, Великого Новгорода и Пскова с отдельными поселениями по Северной Двине, Вятке и Каме.
В течение 1955—1959 годов были обследованы 17 тысяч человек обоих полов из 107 населённых пунктов
На основе данных, собранных Русской антропологической экспедицией, были выделены следующие антропологические типы:
- Ильменско-белозёрский. Головной указатель от 81 до 82, относительно узкое и довольно сильно профилированное лицо. Высокий процент светлых глаз (от 45 до 57), значительный процент светлых оттенков волос (от 29 до 40), сравнительно сильный рост бороды и довольно высокий рост.

Композиционный портрет русских ильменско-белозёрской зоны.
- Валдайский. Отличается высоким головным указателем (82—84), несколько меньшим ростом бороды, более низким и широким лицом. Доля светлых глаз составляет 50 %, волос — около 40 %.

Композиционный портрет русских валдайской зоны.
- Западный верхневолжский. Головной указатель 81. Близок к ильменскому, от которого отличается более тёмной окраской волос (37 % светловолосых), более сильным ростом бороды, более прямой спинкой носа, меньшей частотой складки верхнего века.
- Восточный верхневолжский. Головной указатель 81,4. По сравнению с ильменским и валдайским характеризуется меньшей длиной тела (166 см), более широким лицом, более тёмной окраской глаз и волос. Головной указатель несколько меньше, чем у валдайского, доля вогнутой спинки носа также меньше, чем у валдайского и ильменского.
- Вологдо-вятский. Головной указатель 82,5. Отличается более светлой пигментацией. Процент светлых радужин (без светлосмешанных) — 38, процент светлых волос — 33.

Композиционный портрет русских вологдо-вятской зоны
- Вятско-камский. Головной указатель 81,5. По сравнению с ильменским и валдайским характеризуется меньшей длиной тела, более широким лицом, более тёмной окраской глаз и волос. Головной указатель несколько меньше, чем у валдайского (81,4), доля вогнутой спинки носа также меньше, чем у валдайского и ильменского.
- Клязьминский. Головной указатель 82. Процент светлых радужин (без светлосмешанных) — 38, процент светлых волос — 33. Характерной особенностью типа является сочетание русых волос, карих глаз, брахицефалии, прямых контуров носа, повышенного роста, а также более сильного, чем у ильменского-белозёрского типа, роста бороды (проявляет себя темнопигментированный элемент, не тождественный с другими типами этого региона).
- Центральный. Головной указатель 81,2, процент светлых радужин (без светлосмешанных) — 47, процент светлых волос — 30. Территория однородна антропологически. Средние величины признаков у населения разных районов этой территории очень близки к средним суммарного типа русских. Максимальное сходство с западным верхневолжским.
- Дон-сурский. Головной указатель 79,6. Не имеет аналогов в других группах (сочетание мезокефалии, небольших лицевых размеров, толстогубости и сравнительно сильного роста бороды). От остальных региональных типов юго-восточной территории дон-сурский отличается более светлой пигментацией глаз (частота светлой радужины — 50 %). Понижена частота складки верхнего века.
- Степной. Головной указатель 79,3, процент светлых радужин (без светлосмешанных) — 37 (минимальный по югу), процент светлых волос — 20 (минимальный по региональным типам). По комплексу признаков занимает промежуточное положение между дон-сурским и средневолжским.
- Средневолжский. Головной указатель 80, процент светлых радужин (без светлосмешанных) — 42, процент светлых волос — 21. По комплексу признаков занимает положение, противоположное дон-сурскому (более темная пигментация глаз, больший головной указатель, относительно более низкое лицо, меньший рост бороды).
- Верхнеокский. Головной указатель 81,3, процент светлых радужин (без светлосмешанных) — 47, процент светлых волос — 25. Близок к дон-сурскому, но отличается более высоким головным указателем, относительно более низким лицом, более сильной горизонтальной профилировкой и более темной пигментацией.
- Десно-сейменский. Головной указатель 81,3, процент светлых радужин (без светлосмешанных) — 38, процент светлых волос — 26. Отличается от верхнеокского несколько меньшей высотой лица и большей частотой складки верхнего века.

Композиционный портрет русских десно-сейменской зоны
- Архангельский. Головной указатель 81,5, процент светлых радужин (без светлосмешанных) — 53, процент светлых волос — 45. Очень близок к ильменскому, отличается лишь немного более широким носом, большей частотой светлых глаз, более интенсивным ростом бороды, более профилированным лицом в горизонтальной плоскости и реже встречающейся складкой верхнего века.
- Псковско-поозёрский. Головной указатель 81,8, процент светлых радужин (без светлосмешанных) — 70,5, процент светлых волос — 34,4. Тип выявлен на основе исследований популяции на западном побережье Псковского озера. Население псковского поозёрья по комплексу особенностей относится к западно-балтийскому типу. Обнаруживает сходство с типом балтских племён ятвягов, пруссов и талабов.


На основе этой классификации были выделены и более крупные регионы: северо-запад, северо-восток, юго-запад, юго-восток, в пределах каждого из которых обнаруживается большое единство антропологических типов (5.6).
Одонтология
Морфогенез одонтологических признаков обусловлен главным образом генетическими факторами и слабо зависит от влияния внешней среды. Полиморфизм зубочелюстной системы широко используется в исследованиях проблем этногенеза (7, 8, 9, 10, 11).
Значения основных признаков одонтологии однозначно характеризуют русских как представителей западного круга форм. Одонтологические показатели русского населения наиболее близки к показателям среднеевропейского одонтологического типа.
Межгрупповые различия признаков у русских значительно меньше, чем между русскими и другими народами, что позволяет говорить о единстве русских. Тем не менее, на основании этих различий были выделены 2 основных одонтологических комплекса: северо-западный и южный, связанные между собой рядом переходных форм. Одонтологические типы в составе русского народа напрямую связаны с типами Восточной Европы в целом: среднеевропейским и северным грацильным. Выявлено, что северо-западный тип русских сформировался под влиянием северного грацильного типа Восточной Европы. В частности, для северо-западного одонтологического типа русских характерно некоторое повышение встречаемости лопатообразных резцов, что сближает эти популяции с соседними восточными латышами, эстонцами, финнами, карелами, вепсами и может объясняться влиянием общего субстрата. В целом встречаемость лопатообразных резцов, характерных для монголоидных популяций, у русских повсеместно низка.
Дерматоглифика

Комбинационные полигоны признаков дерматоглифики у русских. Центр окружности соответствует минимальному значению признака по евразийской шкале вариаций, конец радиуса — максимальной величине.
Показано, что дерматоглифические признаки не подвержены влиянию направленного отбора и могут быть использованы в популяционных исследованиях.
Русские популяции являются достаточно однородными по признакам кожного рельефа. Тем не менее, в северных популяциях отмечаются черты северных европеоидов (пониженный дельтовый индекс сочетается с низким процентом добавочных межпальцевых трирадиусов), на юге — южноевропеоидные (относительно высокий процент добавочных межпальцевых трирадиусов сочетается с пониженной частотой осевого проксимального трирадиуса ладони у мужчин и относительно высоким дельтовым индексом у женщин).
Европеоидно-монголоидный комплекс у русских, как правило, невысокий и в среднем составляет 41,9 у мужчин и 44,2 у женщин.
Восточные славяне обнаруживают высокое сходство друг с другом по дерматоглифическим данным. Ключевые признаки кожного рельефа у восточных славян варьируют незначительно, коэффициенты их вариации меньше, чем у европеоидов Европы и на территории Восточной Европы в целом. Дерматоглифический тип восточных славян является характерным для европеоидов Восточной Европы (12, 13, 14, 15).
Генетика
По данным российских генетиков Елены Балановской и Олега Балановского, генофонд русского народа на территории Русской равнины — типично европейский. Процент восточноевразийских генетических маркеров в составе этой выборки не превышает средние показатели по Европе. Результаты изучения этой части русского генофонда показали его близость с населением практически всей Европы, при этом выявлены значительные отличия от населения Урала и Кавказа. Для генофонда выявлена широтная изменчивость (изменение характеристик по оси север — юг).
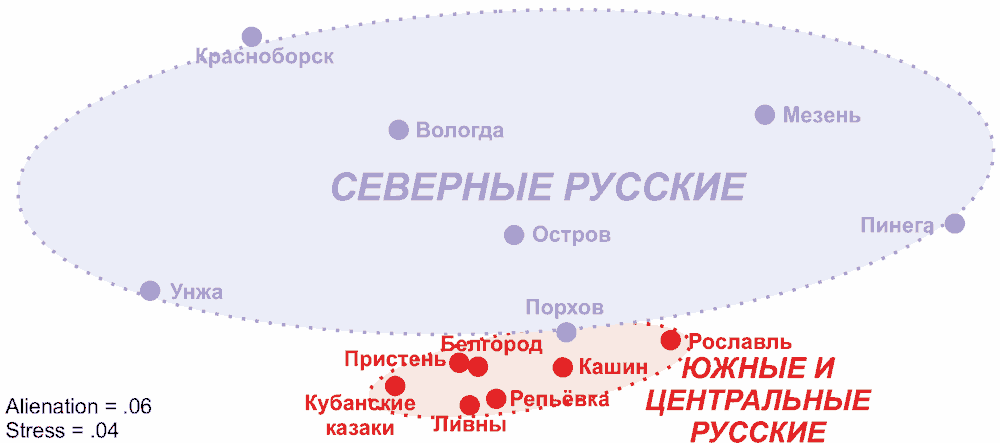
График многомерного шкалирования русских популяций по крупным гаплогруппам Y-хромосомы.
В пределах самого русского генофонда существуют достаточно значительные отличия между северными популяциями русских, и существенно меньшие между южными и центральными, то есть население Русского Севера более гетерогенно.
Последние исследования, включившие в себя комплексный анализ маркеров Y-хромосомы, мтДНК и аутосомных маркеров, позволяют сделать вывод о более выраженном сходстве северных русских с населением Северной Европы в целом. В частности, северные русские по комплексу признаков геногеографии наиболее схожи с балтами (латыши и литовцы) и шведами, достаточно малые генетические расстояния отделяют их от прибалтийско-финских (финны, эстонцы, карелы, вепсы и ижорцы) и некоторых пермско-финскими (коми) народами, а также от многих народов Центральной и Восточной Европы. При этом народы поволжско-финской группы удалены и от всего европейского массива, и от северных русских, что может свидетельствовать об отсутствии предполагавшегося ранее сильного вклада финно-угорского пласта в формирование населения Русского Севера. Таким образом, выявленное сходство генофонда северных русских с генофондами Центральной Европы и севера Восточной Европы позволяет выдвинуть гипотезу о сохранении на этих территориях древнего палеоевропейского генофонда, вошедшего в состав как северных русских, так и остальных народов Северной Европы (16, 17, 18, 19).
Маркеры Y-хромосомы ДНК
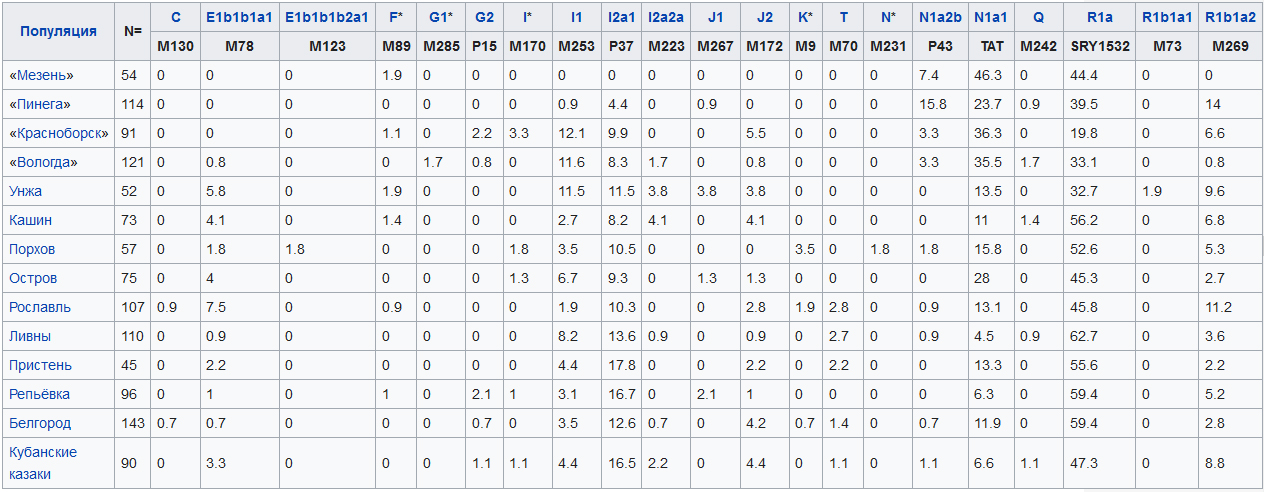
По результатам исследования встречаемости гаплогрупп Y-хромосомы наиболее распространенные вариации гаплогрупп среди русских на 99,7 % совпадают с таковыми из Западной Евразии в рамках по крайней мере последних 3000 лет. Генетический пул русских сходен с остальными популяциями центральной и восточной Европы.
Распределение гаплогрупп Y-хромосомы в некоторых исследованных русских популяциях (19):
Маркеры митохондриальной ДНК
По результатам многомерного анализа частот гаплогрупп митохондриальной ДНК русские входят в общеевропейский кластер популяций, включающий в себя все европейские популяции, за исключением саамов, удмуртов, башкир, ненцев, манси и народов Кавказа (19). Внутри данного кластера можно выделить более узкую группу популяций, в которую входят все европейские народы, в том числе русские, за исключением татар, чувашей и большинства финно-угорских популяций. Гомогенность популяций Европы в пределах данной «узко-европейской» группы высока. Тем не менее, в ней могут быть выделены отдельные кластеры, как правило, совпадающие с этно-лингвистическими группами. Русские входят в «славянский кластер», включающий в себя все исследованные народы славянской группы, а также венгров и эстонцев.
Митохондриальный геном русских в целом типично европейский, доля восточноевразийского компонента в русском митохондриальном геноме не превышает 2 %. Существуют определённые различия в доле восточноевразийского компонента в генофонде различных русских популяций. Среди русских южных и центральных регионов доля восточноевразийских гаплогрупп митохондриальной ДНК составляет 1,3 %, что сравнимо с показателями западных славян (1,6 %), среди северных русских — до 4 %, среди поморов — до 5 % (19).
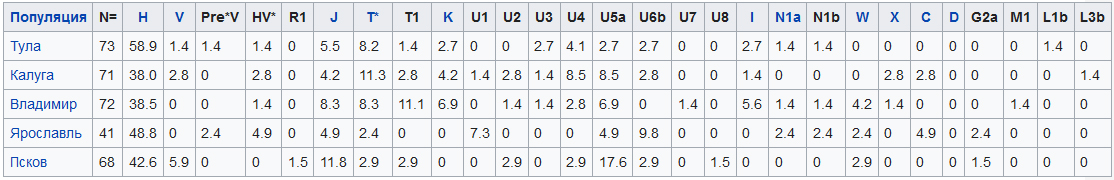
Распределение гаплогрупп митохондриальной ДНК в некоторых исследованных русских популяциях:
Аутосомные маркеры
Аутосомные маркеры — маркеры неполовых хромосом. Как правило, эти гены кодируют синтез определённых белков, ферментов и т. п.
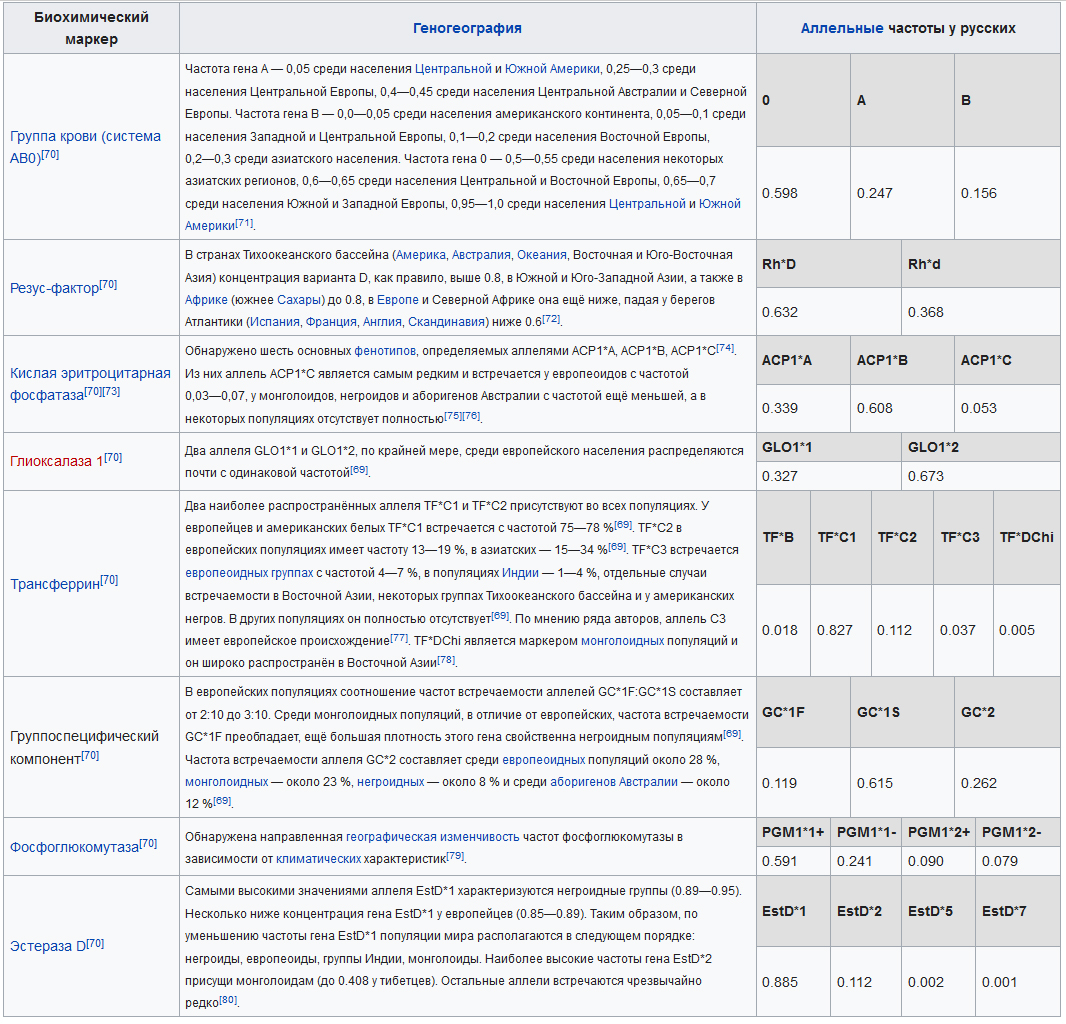
Биохимические маркеры
Генетико-биохимические маркеры, как правило, представляют собой гены, кодирующие синтез ферментов. Полиморфизм некоторых иммунологических маркеров крови также может быть отнесён к биохимическому полиморфизму, так как эти гены кодируют ферменты, катализирующие соединение соответствующих сахаров с группоспецифическими гликопротеидами или гликолипидами. Показано, что полиморфизм многих биохимических маркеров может быть использован в популяционных исследованиях.
В таблице приведены частоты некоторых биохимических маркеров эритроцитарной фракции крови, сыворотки крови, иммунологических маркеров крови (20-31).
Минисателлитные и микросателлитные маркеры
Наиболее изученными в российских популяциях гипервариабельными минисателлитными локусами являются D1S80 , D1S111, IL-1RN и 3’ApoB, так как они представляют наибольший интерес с точки зрения судебно-генетической экспертизы. Изучение полиморфизма локусов D1S80 и 3’ApoB также используется в популяционных исследованиях, так как данные маркеры обладают весьма высокой дифференцирующей способностью. Минисателлитный локус D1S80 локализован на коротком плече 1-й хромосомы. Минисателлитный локус 3’АроВ локализован на коротком плече 2-й хромосомы и представляет собой высокополиморфный участок на расстоянии 500 пар оснований после 3′-конца гена аполипопротеина В.
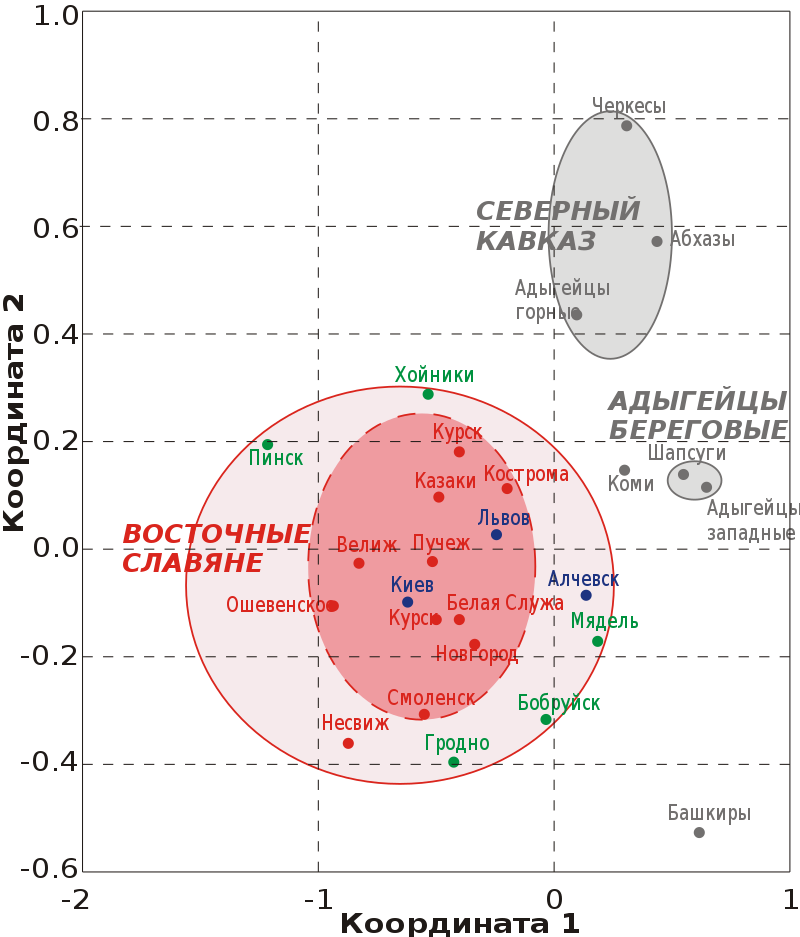
Исследования минисателлитных локусов D1S80 и 3’ApoB показали единство всех восточнославянских популяций и их отличие от коми-зырян, башкир, якутов и северокавказских народов, что может объясняться общим происхождением восточных славян и незначительностью влияния на их генофонд обозначенных этносов.
В 2011 году была исследована генетическая вариабельность 15 широко используемых в судебно-генетической экспертизе микросателлитных маркеров в 17 популяциях Восточной Европы, в том числе в 6 русских городских популяциях (Москва, Белгород, Орёл, Ярославль, Оренбург, Томск.
Все русские группы, а также белорусы, украинцы и коми характеризуются общностью генного пула по указанным маркерам и отсутствием значимой межпопуляционной дифференциации. При этом группа славянских популяций (русские, белорусы, украинцы) значительно отличается от большинства других популяций, в частности, волго-уральского и северокавказского регионов. Все исследованные русские городские популяции, несмотря на значительные географические расстояния между ними, демонстрируют полное отсутствие межпопуляционных различий по исследованным 15 микросателлитным маркерам. По результатам многомерного шкалирования все европейские популяции, включая восточных славян и коми, вошли в общий кластер, что указывает на единство их генофонда. Народы волго-уральского региона и Северного Кавказа образуют отдельные кластеры (32, 33).
Главный комплекс гистосовместимости
Генетические расстояния между популяциями Восточной Европы по результатам исследования полиморфизма 3’ApoB.
Комплекс генов HLA (главного комплекса гистосовместимости) локализован на коротком плече 6-й хромосомы и занимает 3500 тысяч пар оснований, содержит более 220 генов. Он выполняет функции генетического контроля иммунного ответа и поддержания иммунного гомеостаза. Гены локуса HLA DP, DQ и DR кодируют молекулы II класса, выявленные на поверхности антиген-представляющих клеток (хелперов), в частности, В-лимфоцитов, Т-хелперов и некоторых других.
Система HLA является одной из наиболее полиморфных генетических систем. Показано, что, несмотря на влияние направленного естественного отбора, распределение генов HLA II класса адекватно отражает геногеографию этнических групп, и особенности полиморфизма генов HLA могут быть использованы в популяционных исследованиях.
Наибольший полиморфизм проявляет ген DRB1, его изучение наиболее информативно при популяционных исследованиях.
Частоты аллелей гена DRB1 в популяциях восточных славян соответствуют типичному западноевразийскому варианту распределения. Встречаемость частот аллелей DRB1*09 и DRB1*12, характерных для монголоидных популяций, у русских низка.
По результатам многомерного анализа полиморфизма системы HLA в популяциях Европы выделяется три кластера: североевропейский, центральноевропейский и южноевропейский. Русские входят в североевропейский кластер, наряду с немцами, датчанами, голландцами, финнами, ирландцами и англичанами (34-38).
—-
1. Бунак В. В. Происхождение и этническая история русского народа по антропологическим данным. — М.: Наука, 1965. — Т. 88 (новая серия). — (АН СССР. Труды института этнографии им. Н.Н. Миклухо-Маклая).
2. Чебоксаров Н. Н. Монголоидные элементы в населении Центральной Европы // Уч. зап. МГУ. — Москва, 1941. — Вып. 63. — С. 235—270.
3. Дерябин В. Е. Современные восточнославянские народы // Восточные славяне. Антропология и этническая история. — 2002.
4. Алексеев В. П. Происхождение народов Восточной Европы (краниологическое исследование). — М.: Наука, 1969. — 324 с. — 2700 экз.
5. Алексеева Т. И. Этногенез восточных славян по данным антропологии. — М.: Издательство МГУ, 1973.
6. Алексеева Т. И. Антропологический облик русского народа // Русские / В. А. Александров, И. В. Власова, Н. С. Полищук. — М.: Наука, 1999. — 828 с. — (Народы и культуры). — ISBN 5-02-009558-3.
7. Зубов А. А. Этническая одонтология. — М.: Наука, 1973. — 200 с.
8. Зубов А. А., Халдеева Н. И. Одонтология в современной антропологии. — М.: Наука, 1989. — 232 с.
Flower H. W. On the size of the teeth as a character of race (англ.) // J Anthropol. — Institute of Great Britain and Ireland, 1885. — No. 14. — P. 183—186.
9. Ващаева В. Ф. Одонтологическая характеристика русских западных и северо-западных областей РСФСР // Вопросы антропологии. — Издательство Московского университета, 1977а. — Вып. 56. — С. 102—111.
10. Ващаева В. Ф. Одонтологическая характеристика русских центральных, южных и северных областей Европейской части РСФСР // Вопросы антропологии. — Издательство Московского университета, 1977б. — Вып. 57. — С. 133—142.
11. Ващаева В. Ф. Одонтологическая характеристика русского населения Европейской части РСФСР // Вопросы антропологии : Автореферат. — Москва: Издательство Московского университета, 1978.
12. Долинова Н. А. Дерматоглифика восточных славян // Восточные славяне. Антропология и этническая история. — М., 2002.
13. Долинова Н. А. Дерматоглифика русских Европейской части СССР // Проблемы современной антропологии. — Минск, 1983. — С. 43—45.
14. Хить Г. Л. Дерматоглифика народов СССР. — М.: Наука, 1983. — 280 с.
15. Хить Г. Л., Долинова Н. А. Расовая дифференциация человечества (дерматоглифические данные). — М.: Наука, 1990. — 201 с.
16. Елена Балановская, Олег Балановский. Русский генофонд. Генетика русских. Лаборатория популяционной генетики человека Медико-генетического научного центра РАМН.
17. Елена Балановская, Олег Балановский. Русский генофонд. Генетика русских Архивная копия от 15 февраля 2015 на Wayback Machine Лаборатория популяционной генетики человека Медико-генетического научного центра РАМН. 2007.
18. Балановская Е. В., Пежемский Д. В., Романов А. Г., Баранова Е. Е., Ромашкина М. В., Агджоян А. Т., Балаганский А. Г., Евсеева И. В., Виллемс Р., Балановский О. П. Генофонд Русского Севера: славяне? Финны? Палеоевропейцы // Антропология. — Вестник Московского университета. Серия XXIII, 2011. — Вып. 3.
19. Балановский О. П. Изменчивость генофонда в пространстве и времени синтез данных о геногеографии митохондриальной ДНК и Y-хромосомы, 2012.
20.(69 в таблице)Прокоп О., Гелер В. Группы крови человека / О. Прокоп, В. Гелер; Пер. с нем. А. С. Гладких; Под ред. В. В. Томилина. — М.: Медицина, 1991. — 510, [1] с. — ISBN 5-225-00541-1.
21. (70 в таблице) База данных «Частоты аутосомных маркеров в популяциях Евразии» Архивная копия от 4 марта 2016 на Wayback Machine Версия 1.0. © Почешхова Э. А., Соловьёва Д. С., Балановская Е. В. Лаборатория популяционной генетики человека, ГУ МГНЦ РАМН. 2007.
22. (71) Mourant A. E., Kopec A. C., Domaniewska-Sobesak K. The Distribution of the Human Blood Groups and other Polymorphism. — London: Oxford University Press, 1976.
23.(72) Чебоксаров Н. Н., Чебоксарова И. А. Народы, расы, культуры. — 2-е изд., испр. и доп. — М.: Наука, 1985. — 272 с.
24. (73) Ирисова О. В. Полиморфизм эритроцитарной кислой фосфатазы в различных группах населения Советского Союза // Вопросы антропологии. — 1976. — Вып. 53.
25. (74) Hopkinson D. A., Spenser N., Harris H. Red cell asid phosphatase variants: a new human polymorphism (англ.) // Nature. — London, 1963. — No. 199. — P. 969—974.
26. (75) Spenser N., Hopkinson D. A., Harris H. Quantitative differences and gene dosage in the human red cell asid phosphotase polymorphism (англ.) // Nature. — London, 1964. — No. 201. — P. 748—753.
27. (76) Спицын В. А., Афанасьева И. С., Ирисова О. В., Боева С. Б. О взаимосвязи между климатическими факторами и географическим распределением генов эритроцитарной кислой фосфатазы человека // Науч. докл. высш. школы. Биол. фак. МГУ. — 1977. — Вып. 3. — С. 68—73.
28. (77) Sikstrom C., Nylander P. O. Transferrin C subtypes and ethnic heterogeneity in Sweden (англ.) // Hum. Hered. — 1990. — Vol. 40. — P. 335—339.
29. (78) Kamboh M. I., Ferrell R. E. Human transferrin polymorphism (англ.) // Hum. Hered. — 1987. — Vol. 37. — P. 65—81.
30. (79) Спицын В. А. Биохимический полиморфизм человека. — М.: Издательство Московского университета, 1985. — 216 с.
31. (80) Papiha S. S., Nahar A. The world distribution of the electrophoretic variants of the red cell enzyme Esterase D (англ.) // Hum. Hered. — 1987. — Vol. 27. — P. 424—432.
32. Dmitry A. Verbenko, A. N. Knjazev, A. I. Mikulich, E. K. Khusnutdinova, N. A. Bebyakova, S. A. Limborska. Variability of the 3’ApoB Minisatellite Locus in Eastern Slavonic Populations (англ.) // Hum Hered. — 2005.
33. Степанов В. А. и др. Характеристика популяций Российской Федерации по панели пятнадцати локусов, используемых для ДНК-идентификации и в судебно-медицинской экспертизе // Acta Naturae. — 2011. — Т. 3, № 2. — С. 59—71.
34. James Robinson, Matthew J. Waller, Peter Parham, Julia G. Bodmer and Steven G. E. Marsh. IMGT/HLA Database — a sequence database for the human major histocompatibility complex (англ.) // Nucleic Acids Research. — 2001. — Vol. 29, no. 1. — P. 210—213.
35. Хаитов Р. М., Алексеев Л. П. Система генов HLA и регуляция иммунного ответа // Аллергия, астма и клиническая иммунология. — 2000. — № 8. — С. 7—16.
36. Ярилин А. А. Основы иммунологии. — М.: Медицина, 1999. — С. 52—53, 223—225. — 608 с. — (Учебная литература для студентов медицинских вузов). — 10 000 экз. — ISBN 5-225-02755-5.
37. Хромова Н. А. Полиморфизм системы HLA у представителей разных славянских этнических групп (русской, белорусской и украинской) : Автореферат. — Москва, 2006.
38. Болдырева М. Н. HLA (класс II) и естественный отбор. «Функциональный» генотип, гипотеза преимущества «функциональной» гетерозиготности : Автореферат. — Москва, 2007. — С. 9—16.
Читайте также:
• Происхождение русских и других славян по данным генетики
